
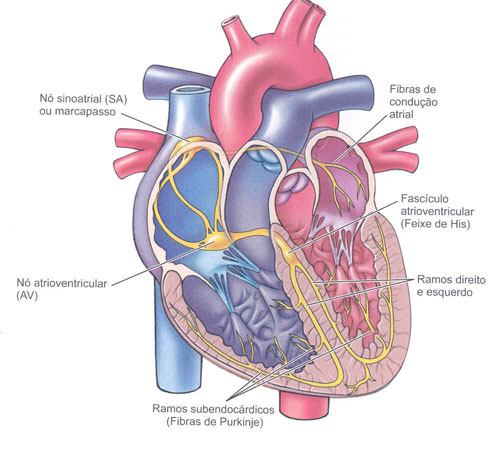
O coração é um órgão único e muscular que
possui duas características que o distingue dos demais órgãos, a excitabilidade
e a condutibilidade. Este é capaz de gerar estímulos espontâneos de maneira rítmica,
sem necessitar de inervação parar gerar um estímulo elétrico para sua
contração. O nó sinoatrial (S-A), as vias internodais, o nó atrioventricular
(A-V), o feixe atrioventricular e as fibras de Purkinje fazem parte desse
sistema.
Fonte: http://dc220.4shared.com/doc/6WX5x04b/preview_html_m111e5721.jpg
Os músculos cardíacos apresentam três
tipos de canais iônicos, os quais são importantes para a variação da voltagem
da membrana que é necessária para a geração do potencial de ação cardíaco. O
potencial de membrana da fibra muscular é de aproximadamente -90mV e quando um
impulso que induz a despolarização chega até ela, acontece as seguintes etapas:
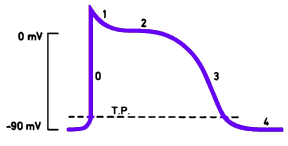
0
- Nesta fase há a abertura dos canais rápidos de Na+, os quais vão elevar o
potencial de membrana (despolarizar).
1
– Abertura dos canais de K+, os quais vão iniciar a repolarização da célula.
2
– Nesta etapa há a abertura dos canais lentos de Ca2+, que começam a se abrir
lentamente até abrir por completo. Esses canais permitem o influxo do íon Ca2+
da célula, o que vai interromper a queda do potencial. O influxo de Ca2+ “compensa”
o efluxo de K+ da célula, o que vai retardar a repolarização, formando o
“platô”.
3
– Canais lentos de Ca2+ se fecham. A saída contínua de K+ leva a repolarização
da célula e, consequentemente, faz com que esta chegue ao seu potencial de
repouso.
4
– Canais de K+ se fecham e membrana se mantém em seu potencial de repouso.
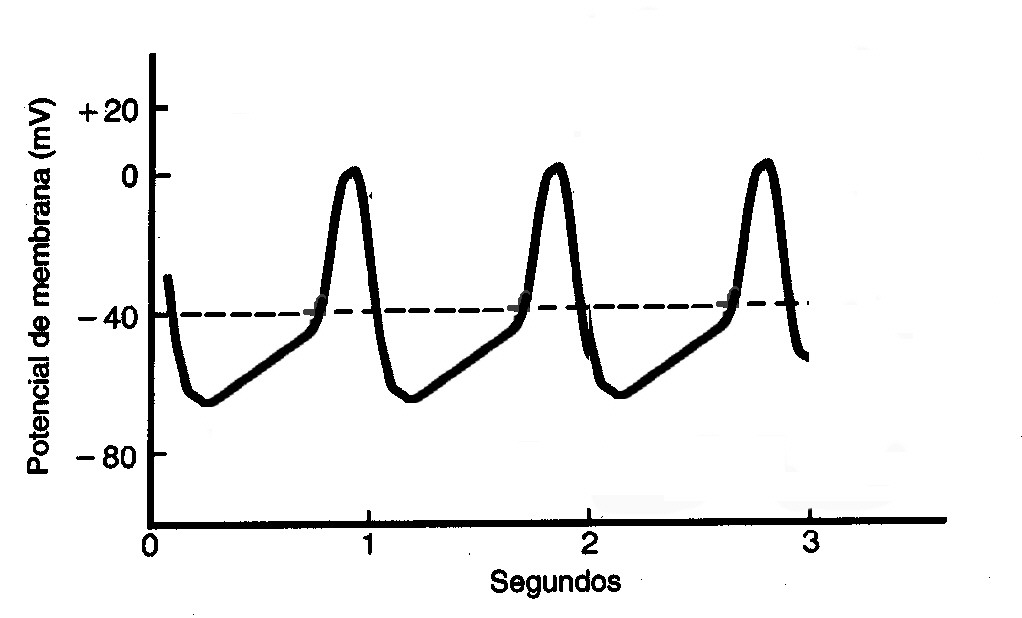
O potencial de ação do nó sinoatrial e
atrioventricular se difere do potencial da fibra cardíaca. Nestes o potencial
de repouso é mais negativo e os canais lentos de Na+ ficam inativos (não podem
ser abertos). Além disso, o potencial de ação vai ter início com a
despolarização causada pela abertura dos canais lentos de Na+. Dessa forma, o
potencial vai ser caracterizado por um desenvolvimento mais lento e decréscimo
menos acentuado.
A fase 4 destes é marcada por uma
despolarização espontânea (influxo constante de Na+ na célula), uniforme e
lenta. Dessa forma, a célula tem uma despolarização espontânea até chegar ao
potencial limiar e induzir o potencial de ação.
Como demonstrado na figura, o nó
sinoatrial possui uma maior freqüência de disparo do potencial de ação e, por
este motivo, ele é o marcapasso do coração. Como a despolarização do nó
sinoatrial é mais rápida, seu impulso é gerado e conduzido por meio do átrio
até o nó atrioventricular. Este que ainda não despolarizou o suficiente até
gerar o potencial de ação recebe o impulso vindo do nó sinoatrial e atinge seu
limiar (ocorre o potencial de ação) e transmite o impulso elétrico aos
ventrículos.
Potencial de ação no nó Sinoatrial
Potencial de ação no nó Atrioventricular
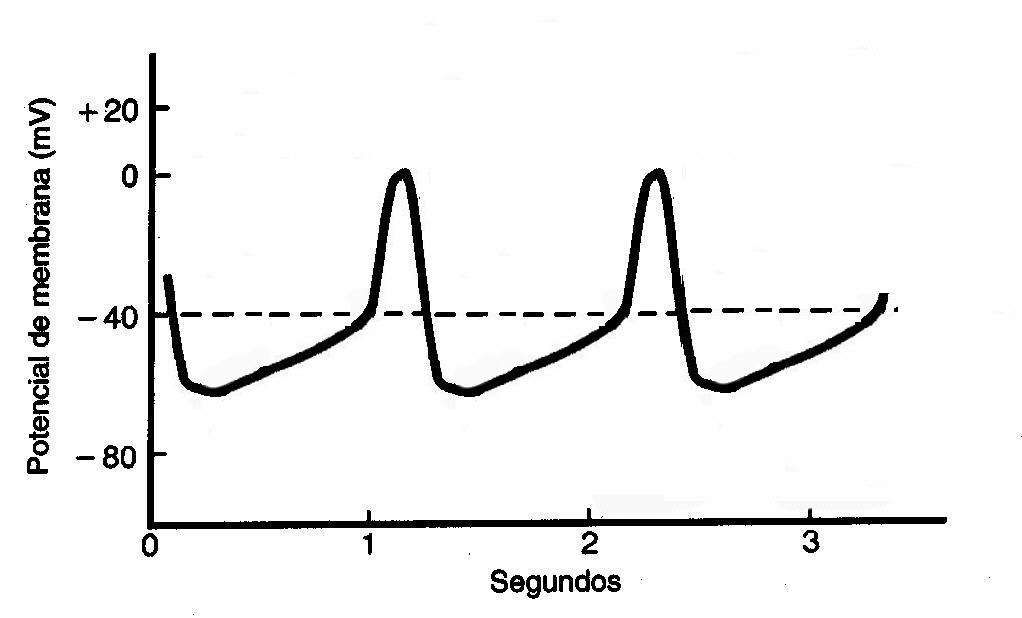
O nó atrioventricular é importante
para o retardo da transmissão do impulso elétrico do átrio para o ventrículo,
fazendo assim com que os átrios se contraiam antes dos ventrículos.
A
CONTRAÇÃO CARDÍACA
O sarcômero é a
unidade contrátil do músculo, neste se encontra filamentos grossos e finos que
se intercalam. O filamento fino possui actina, troponina e tropomiosina. A
actina é uma proteína que quando polimeralizada forma uma dupla hélice que irá
conter os sítios de ligação com a miosina. A tropomiosina está ligada a actina
e tem função de impedir a ligação desta com a miosina (bloqueia o sitio de
ligação). Por fim, a proteína troponina fica ligada a tropomiosina e tem como
função regular o bloqueio feito pela tropomiosina. Além disso, essa proteína
possui três regiões: uma com afinidade a actina, outro com a tropomiosina e a
última ao Ca2+.

Fonte: http://www.apoioescolar24horas.com.br/cf/salaaula/estudos/biologia/054_histologia/img/m2.jpg
Os
filamentos de actina e miosina ligam-se facilmente quando não estão sob
bloqueio da tropomiosina (que impede a ligação na ausência de Ca2+). O sitio de
ligação da actina/miosina é liberado com a chegada do potencial de ação na
fibra muscular, que vai permitir a entrada de Ca2+ na célula (liberados pelo
reticulo endoplasmático). Este se liga a troponina C, a qual vai sofrer uma
mudança conformacional e fazer com que a tropomiosina libere os sítios de
ligação da actina com a miosina.
A
interação entre actina e miosina só acontece com a presença de ATP e magnésio
(estão presentes em condições normais).
A
contração irá acontecer com o deslizamento dos filamentos finos sobre os grossos
(encurtamento do sarcômero), com as seguintes etapas:
- A molécula de ATP se liga a um sítio presente na cabeça da miosina e é hidrolisada em ADP e Pi, que vão permaner fixos a cabeça, ocupando o sítio. Nesse estado, a cabeça pode se entender em direção ao filamento fino.
- Com a liberação do sítio actina/miosina (bloqueada pelo complexo troponina-tropomiosina) proporcionada pela ligação do Ca2+ à troponina C, os filamentos de actina e miosina podem se ligar.
- A miosina diminui sua afinidade com o ADP e Pi, os quais vão se dissociar do sítio catalítico da miosina. Simultaneamente a dissociação, a cabeça da miosina se move e puxa o filamento de actina, provocando seu deslizamento sobre o filamento de miosina.
- Para a dissociação da miosina a actina, uma molécula de ATP tem que se ligar novamente a ela. Com a entrada de um ATP a molécula retorna a sua conformação original e promove a quebra do ATP, o que leva ao recomeço do ciclo.

Nenhum comentário:
Postar um comentário